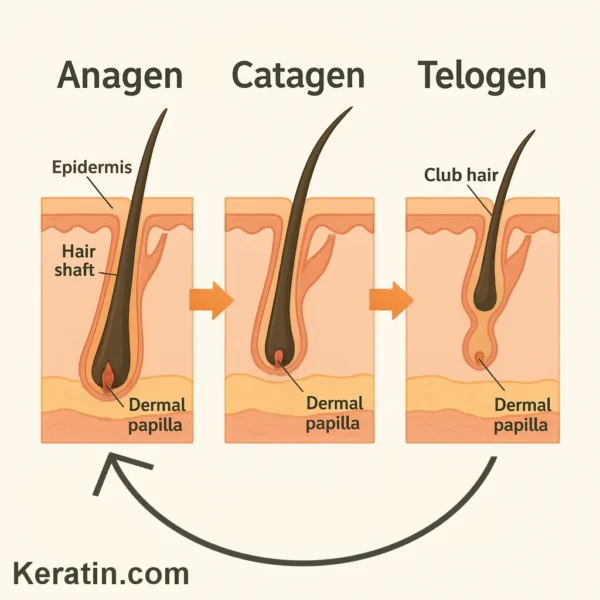
Human scalp hair does not grow continuously from birth to old age. Instead, every hair follicle runs through a repeating cycle of growth, regression, and rest. At any given moment, neighbouring follicles are usually at different stages, which is why people do not shed all their hair at once. Appreciating this rhythm helps to explain why hair length varies on different body sites, why daily shedding is normal, and why some medical conditions or treatments lead to thinning or loss.
The architecture of a growing follicle: A hair follicle is a tiny, complex organ sunk deep in the skin. In its fully active state it contains multiple specialised cell layers arranged like sleeves around the growing hair fiber. In mature growth, scientists describe eight main concentric layers, each performing tasks such as driving cell division, shaping the hair shaft, forming its protective cuticle, or supplying pigment. Melanocytes, the pigment‑forming cells, reside next to the developing fiber and add melanin during growth; that is why color is laid down only while the hair is actively forming. When growth pauses, pigment production pauses too, and new grey hairs emerge when pigment cells become depleted or inactive. Check out a page on hair follicle structure here.
Phases of the cycle: anagen, catagen and telogen: The classic human hair cycle has three main phases.
Anagen: active growth: Anagen is the longest and most industrious phase, during which the hair fiber is produced. On a healthy scalp, roughly 85–90% of follicles are in anagen at any time. Within anagen, researchers distinguish early, middle and late sub‑stages – proanagen, mesanagen and metanagen – that describe the rapid build‑up from initiation to full power. Proanagen marks the restart of the machinery: DNA and RNA synthesis surge, the matrix cells at the base begin dividing, and the follicle elongates downwards into the dermis. Through mesanagen the follicle expands in length and girth, and by metanagen it reaches maximum size, with robust proliferation and differentiation producing a thick, pigmented hair. How fast does an anagen hair grow? On the scalp, average growth is about 0.35 millimetres per day – roughly one centimetre per month – though this varies with age, sex, ethnicity, health, and body site. The length a hair can achieve is governed not by its daily growth rate but by how long an individual follicle remains in anagen before it switches off. For scalp follicles, anagen typically lasts about three to six years. Someone whose follicles commonly remain at the upper end of that range can grow very long hair; someone with shorter anagen spans will find their hair reaches a natural limit sooner even if it is healthy.Catagen, controlled regression: Anagen does not end abruptly. The follicle transitions through catagen, a brief but highly choreographed winding‑down phase lasting about two to three weeks. During catagen, cell division stops, the lower part of the follicle shrinks dramatically, and the hair bulb – where growth previously occurred – undergoes programmed regression. Importantly, this is not damage but a normal, actively regulated process. Structures that are no longer needed are dismantled and recycled. The follicle moves its regenerative stem cell populations into a protected niche near the insertion of the tiny arrector pili muscle, ready for the next cycle.Telogen, resting and readiness: Following catagen the follicle enters telogen, often called the “resting” phase. The hair produced in the previous cycle remains anchored near the skin surface, but the follicle itself is mostly dormant. On the scalp, telogen usually lasts around 30–90 days. At any moment about 10% of scalp follicles are in telogen. Telogen is not idle in a biological sense: the follicle is primed to re‑enter proanagen, and signalling between the follicle, surrounding skin, nerves and immune cells helps determine when the next growth burst will begin.Exogen and kenogen: Many people notice daily hair shedding during washing or brushing. This is largely due to the timing of telogen hairs releasing from their follicles. Some researchers recognise a distinct shedding phase, called exogen , to describe the time point at which the old fiber actually lets go. Another term you may encounter is kenogen , a lag period in which the follicle is empty between shedding the old hair and starting a new one. These refinements do not change the core concept: each follicle alternates between growing a fiber, regressing, resting, and then starting again. Check out a page that discusses the changes in hair cycling during androgenetic alopecia . Follicles dance to their own beat: In some mammals, body hair cycles in visible waves across the skin. Human follicles are different: they tend to cycle independently. That independence is a protective advantage – if all scalp hairs rested at once, we would shed our coverage in a molt. Instead, the mosaic pattern ensures overall density is maintained, even though individual hairs are constantly being renewed. Subtle coordination does exist, however. Local chemical signals, the body’s hormones, nutrition, stress mediators and immune factors can nudge groups of follicles toward growth or rest together, which becomes apparent during certain life events or illnesses. Check out another page on seasonal hair shedding .
What influences the cycle: Because the cycle depends on living cells that divide, differentiate and respond to signals, many internal and external factors can lengthen or shorten phases.
Ageing. With advancing age, anagen duration tends to shorten and hair fibers miniaturise, producing finer, shorter hairs. Scalp density can slowly decline as a result.Genetics and hormones. In androgenetic alopecia (often called male‑ or female‑pattern hair loss) genetically susceptible follicles respond to androgens by shortening anagen and producing progressively thinner fibers. Over many cycles this “miniaturisation” leads to visible thinning.The immune system. In alopecia areata, immune cells attack the anagen hair bulb, abruptly terminating growth and pushing follicles into an early rest state; regrowth is often possible because the stem cell reservoir remains.Medications and toxins. Drugs that target rapidly dividing cells, such as some chemotherapies, can cause anagen effluvium , a sudden loss of growing hairs because matrix cells cannot sustain fiber production. Other medicines, major surgery, fever, or significant life stress can trigger telogen effluvium , in which many follicles synchronously enter telogen and increased shedding follows about two to three months later. Because telogen is temporary, shedding usually settles as the next anagen wave begins.Radiation, scarring and tumours. Physical destruction of the follicle, through burns, radiation therapy, scarring skin diseases or tumour infiltration, can abolish its ability to cycle. In scarring alopecias the follicular stem cell niche is replaced by fibrous tissue; early recognition and treatment aim to halt further loss.Cosmetics and grooming. Shampoos, dyes and styling do not alter the timing of follicle phases deep in the skin, although harsh treatments can break fibers and increase the impression of shedding.Nutrition and general health. Severe iron deficiency, extreme dieting, thyroid disorders and systemic illness can all disturb the cycle. Correcting the underlying issue usually allows follicles to re‑enter anagen. Timing explains everyday observations: Understanding durations helps make sense of common experiences. If telogen on the scalp lasts about 1–3 months, a stressful event in the summer time may not be followed by noticeable shedding until early autumn, for example. Likewise, if anagen lasts several years, it takes time to rebuild density after a widespread telogen effluvium: the new hairs are present, but they need months to grow long enough to contribute meaningfully to volume. People often ask whether cutting hair makes it grow faster. Trimming does not change the follicle’s internal clock; it simply removes split ends and can make hair look fuller while the follicles continue at their programmed pace under the skin.
From cycle to clinic, what doctors look for: When clinicians evaluate hair loss, they consider which phase is being disrupted. Increased club hairs (the tapered hairs shed at the end of telogen) suggest a shift toward telogen, as in telogen effluvium. Broken, tapered or pigment‑depleted hairs can point to immune‑mediated interruption of anagen, as in alopecia areata. In pattern hair loss, serial photographs and trichoscopy reveal a mix of thick and miniaturised hairs, reflecting shortened anagen spans and reduced follicle caliber. Treatments are chosen to prolong anagen, enlarge miniaturised follicles, quieten inflammation, or remove the trigger that pushed follicles into rest.
Why pigment and growth are linked: Because melanogenesis, the making of melanin, occurs only while the fiber is being constructed, colour is tightly coupled to anagen. Grey or white hairs appear when pigment cells are no longer delivering melanin during growth. This can happen gradually with age as melanocyte reservoirs deplete, or more abruptly during certain immune conditions. Once the follicle enters catagen and telogen, pigment production stops; a new cycle must begin before colour can return.
A lifelong rhythm: From infancy to old age, each human hair follicle repeats this choreography: build a hair, stand down, pause, and start again. Most follicles persist for life, even if the fibers they generate change in thickness, colour or length. The remarkable resilience of this system underlies newer therapeutic strategies that aim not merely to replace hairs but to encourage follicles back into healthy anagen and maintain them there.
Key takeaways: There are a few take away facts from research published on the human hair growth cycle.
Every scalp follicle cycles through growth (anagen), controlled regression (catagen) and rest (telogen), with shedding (exogen) marking release of the old fiber. About 85–90% of scalp follicles are in anagen at any time; around 10% are in telogen. Scalp hair grows roughly 0.35 mm per day. Anagen commonly lasts 3–6 years, catagen about 2–3 weeks, and telogen about 1–3 months. The length a hair can reach is mostly determined by the time duration of an individual follicle’s anagen period, not the monthly growth rate. Follicles cycle largely independently of each other, creating a mosaic that maintains overall coverage. Genetics, hormones, immune activity, medications, illness, scarring and ageing can all shift the balance between phases, leading to shedding or thinning. Because phase changes take time, the effects of a trigger may appear months later, and recovery likewise unfolds over months to years as new anagen hairs lengthen. Further reading: For readers who wish to explore the science in more depth, classic and modern reviews include work by Saitoh and colleagues on the human cycle, Moretti et al. on re‑evaluating its timing, Kligman’s analyses of age‑related changes, and more contemporary guides such as Oh et al. that describe how researchers study hair growth cycling in vivo. These and other sources expand on the cellular choreography and the molecular “clocks” that keep our follicles in step, even as they dance to their own beat.
Bibliography
11711645 {11711645:4WMX33EI},{11711645:EMR7Y9HA},{11711645:BIKNPAS9},{11711645:FYHQZMUH},{11711645:FRHNDBFD},{11711645:SP5A8BKW},{11711645:6CR2WRWS},{11711645:PFFVSYJG},{11711645:I2Z4MZNU},{11711645:I93UBY9X},{11711645:77577G83},{11711645:QCVYSNPB},{11711645:4NLSWUL5},{11711645:CU9GUKS6},{11711645:2X8WCSQE},{11711645:SJYUSA3C},{11711645:8QSSP5HH},{11711645:KWN3H35S},{11711645:LJ5F2RFC} 1 vancouver 50 date asc 1909 https://www.keratin.com/wp-content/plugins/zotpress/ %7B%22status%22%3A%22success%22%2C%22updateneeded%22%3Afalse%2C%22instance%22%3Afalse%2C%22meta%22%3A%7B%22request_last%22%3A0%2C%22request_next%22%3A0%2C%22used_cache%22%3Atrue%7D%2C%22data%22%3A%5B%7B%22key%22%3A%22QCVYSNPB%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Chase%20et%20al.%22%2C%22parsedDate%22%3A%221953-05%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BChase%20HB%2C%20Montagna%20W%2C%20Malone%20JD.%20Changes%20in%20the%20skin%20in%20relation%20to%20the%20hair%20growth%20cycle.%20Anat%20Rec.%201953%20May%3B116%281%29%3A75%26%23x2013%3B81.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Changes%20in%20the%20skin%20in%20relation%20to%20the%20hair%20growth%20cycle%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22H.%20B.%22%2C%22lastName%22%3A%22Chase%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22W.%22%2C%22lastName%22%3A%22Montagna%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22J.%20D.%22%2C%22lastName%22%3A%22Malone%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22date%22%3A%221953-05%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1002%5C%2Far.1091160107%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220003-276X%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A20%3A36Z%22%7D%7D%2C%7B%22key%22%3A%224NLSWUL5%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Chase%22%2C%22parsedDate%22%3A%221954-01%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BChase%20HB.%20Growth%20of%20the%20hair.%20Physiol%20Rev.%201954%20Jan%3B34%281%29%3A113%26%23x2013%3B26.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Growth%20of%20the%20hair%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22H.%20B.%22%2C%22lastName%22%3A%22Chase%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22date%22%3A%221954-01%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1152%5C%2Fphysrev.1954.34.1.113%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220031-9333%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A20%3A07Z%22%7D%7D%2C%7B%22key%22%3A%222X8WCSQE%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Van%20Scott%20et%20al.%22%2C%22parsedDate%22%3A%221957-09%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BVan%20Scott%20EJ%2C%20Reinertson%20RP%2C%20Steinmuller%20R.%20The%20growing%20hair%20roots%20of%20the%20human%20scalp%20and%20morphologic%20changes%20therein%20following%20amethopterin%20therapy.%20J%20Invest%20Dermatol.%201957%20Sept%3B29%283%29%3A197%26%23x2013%3B204.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22The%20growing%20hair%20roots%20of%20the%20human%20scalp%20and%20morphologic%20changes%20therein%20following%20amethopterin%20therapy%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22E.%20J.%22%2C%22lastName%22%3A%22Van%20Scott%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22R.%20P.%22%2C%22lastName%22%3A%22Reinertson%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22R.%22%2C%22lastName%22%3A%22Steinmuller%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22date%22%3A%221957-09%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1038%5C%2Fjid.1957.87%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220022-202X%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A11%3A55Z%22%7D%7D%2C%7B%22key%22%3A%22CU9GUKS6%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Pelfini%20et%20al.%22%2C%22parsedDate%22%3A%221969%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BPelfini%20C%2C%20Cerimele%20D%2C%20Pisanu%20G.%20Aging%20of%20the%20skin%20and%20hair%20growth%20in%20man.%20In%3A%20Hair%20growth.%20London%3A%20Pergamon%20Press%3B%201969.%20p.%20153%26%23x2013%3B60.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22bookSection%22%2C%22title%22%3A%22Aging%20of%20the%20skin%20and%20hair%20growth%20in%20man%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22C%22%2C%22lastName%22%3A%22Pelfini%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22D%22%2C%22lastName%22%3A%22Cerimele%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22G%22%2C%22lastName%22%3A%22Pisanu%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22bookTitle%22%3A%22Hair%20growth%22%2C%22date%22%3A%221969%22%2C%22originalDate%22%3A%22%22%2C%22originalPublisher%22%3A%22%22%2C%22originalPlace%22%3A%22%22%2C%22format%22%3A%22%22%2C%22ISBN%22%3A%22%22%2C%22DOI%22%3A%22%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22ISSN%22%3A%22%22%2C%22language%22%3A%22%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A15%3A29Z%22%7D%7D%2C%7B%22key%22%3A%22KWN3H35S%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Saitoh%20et%20al.%22%2C%22parsedDate%22%3A%221970-01%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BSaitoh%20M%2C%20Uzuka%20M%2C%20Sakamoto%20M.%20Human%20hair%20cycle.%20J%20Invest%20Dermatol.%201970%20Jan%3B54%281%29%3A65%26%23x2013%3B81.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Human%20hair%20cycle%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22M.%22%2C%22lastName%22%3A%22Saitoh%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22M.%22%2C%22lastName%22%3A%22Uzuka%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22M.%22%2C%22lastName%22%3A%22Sakamoto%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22date%22%3A%221970-01%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1111%5C%2F1523-1747.ep12551679%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220022-202X%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A08%3A21Z%22%7D%7D%2C%7B%22key%22%3A%22LJ5F2RFC%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Moretti%20et%20al.%22%2C%22parsedDate%22%3A%221976-05%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BMoretti%20G%2C%20Rampini%20E%2C%20Rebora%20A.%20The%20hair%20cycle%20re-evaluated.%20Int%20J%20Dermatol.%201976%20May%3B15%284%29%3A277%26%23x2013%3B85.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22The%20hair%20cycle%20re-evaluated%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22G.%22%2C%22lastName%22%3A%22Moretti%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22E.%22%2C%22lastName%22%3A%22Rampini%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22A.%22%2C%22lastName%22%3A%22Rebora%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22date%22%3A%221976-05%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1111%5C%2Fj.1365-4362.1976.tb00701.x%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220011-9059%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A07%3A18Z%22%7D%7D%2C%7B%22key%22%3A%228QSSP5HH%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Ebling%22%2C%22parsedDate%22%3A%221988%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BEbling%20FJ.%20The%20hair%20cycle%20and%20its%20regulation.%20Clin%20Dermatol.%201988%3B6%284%29%3A67%26%23x2013%3B73.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22The%20hair%20cycle%20and%20its%20regulation%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22F.%20J.%22%2C%22lastName%22%3A%22Ebling%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22date%22%3A%221988%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1016%5C%2F0738-081x%2888%2990068-5%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220738-081X%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A08%3A33Z%22%7D%7D%2C%7B%22key%22%3A%22SJYUSA3C%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Kligman%22%2C%22parsedDate%22%3A%221988%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BKligman%20AM.%20The%20comparative%20histopathology%20of%20male-pattern%20baldness%20and%20senescent%20baldness.%20Clin%20Dermatol.%201988%3B6%284%29%3A108%26%23x2013%3B18.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22The%20comparative%20histopathology%20of%20male-pattern%20baldness%20and%20senescent%20baldness%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22A.%20M.%22%2C%22lastName%22%3A%22Kligman%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22date%22%3A%221988%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1016%5C%2F0738-081x%2888%2990074-0%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220738-081X%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A10%3A42Z%22%7D%7D%2C%7B%22key%22%3A%22FRHNDBFD%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Courtois%20et%20al.%22%2C%22parsedDate%22%3A%221994%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BCourtois%20M%2C%20Loussouarn%20G%2C%20Hourseau%20C%2C%20Grollier%20JF.%20Hair%20cycle%20and%20alopecia.%20Skin%20Pharmacol.%201994%3B7%281%26%23x2013%3B2%29%3A84%26%23x2013%3B9.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Hair%20cycle%20and%20alopecia%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22M.%22%2C%22lastName%22%3A%22Courtois%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22G.%22%2C%22lastName%22%3A%22Loussouarn%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22C.%22%2C%22lastName%22%3A%22Hourseau%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22J.%20F.%22%2C%22lastName%22%3A%22Grollier%22%7D%5D%2C%22abstractNote%22%3A%22Male%20pattern%20alopecia%20is%20the%20outcome%20of%20profound%20modifications%20in%20the%20duration%2C%20succession%20and%20frequency%20of%20hair%20cycles.%20These%20phenomena%20were%20studied%20by%20phototrichogram%20in%2010%20male%20subjects%2C%20with%20or%20without%20alopecia%2C%20over%20a%20period%20of%2015%20years.%20Almost%2010%2C000%20hair%20cycles%20were%20accounted%20for%2C%20yielding%20a%20detailed%20picture%20of%20the%20alopecia%20condition%3A%20%281%29%20A%20decrease%20in%20the%20duration%20of%20anagen%20for%20a%20certain%20proportion%20of%20hairs%2C%20a%20proportion%20which%20increases%20in%20size%2C%20the%20more%20advanced%20the%20alopecia%3B%20the%20result%20of%20this%20premature%20transformation%20from%20anagen%20to%20telogen%20is%20an%20increase%20in%20the%20rate%20of%20hair%20loss.%20%282%29%20A%20parallel%20decline%20in%20hair%20diameter.%20%283%29%20Longer%20latency%20periods%20between%20the%20fall%20of%20a%20hair%20and%20the%20onset%20of%20regrowth%2C%20leading%20to%20a%20reduction%20in%20the%20number%20of%20hairs%20present%20on%20the%20scalp%20surface.%20The%20shorter%20finer%20hairs%20are%20absent%20more%20frequently%20and%20absent%20for%20longer%20periods%20and%20this%20contributes%20to%20the%20effect%20of%20alopecia.%22%2C%22date%22%3A%221994%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1159%5C%2F000211279%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%221011-0283%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A37%3A14Z%22%7D%7D%2C%7B%22key%22%3A%22FYHQZMUH%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Paus%20and%20Cotsarelis%22%2C%22parsedDate%22%3A%221999-08-12%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BPaus%20R%2C%20Cotsarelis%20G.%20The%20biology%20of%20hair%20follicles.%20N%20Engl%20J%20Med.%201999%20Aug%2012%3B341%287%29%3A491%26%23x2013%3B7.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22The%20biology%20of%20hair%20follicles%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22R.%22%2C%22lastName%22%3A%22Paus%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22G.%22%2C%22lastName%22%3A%22Cotsarelis%22%7D%5D%2C%22abstractNote%22%3A%22%22%2C%22date%22%3A%221999-08-12%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1056%5C%2FNEJM199908123410706%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220028-4793%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A37%3A25Z%22%7D%7D%2C%7B%22key%22%3A%2277577G83%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Paus%20et%20al.%22%2C%22parsedDate%22%3A%221999-12%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BPaus%20R%2C%20M%26%23xFC%3Bller-R%26%23xF6%3Bver%20S%2C%20Botchkarev%20VA.%20Chronobiology%20of%20the%20hair%20follicle%3A%20hunting%20the%20%26%23x201C%3B%20hair%20cycle%20clock.%26%23x201D%3B%20J%20Investig%20Dermatol%20Symp%20Proc.%201999%20Dec%3B4%283%29%3A338%26%23x2013%3B45.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Chronobiology%20of%20the%20hair%20follicle%3A%20hunting%20the%20%5C%22%20hair%20cycle%20clock%5C%22%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22R.%22%2C%22lastName%22%3A%22Paus%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22S.%22%2C%22lastName%22%3A%22M%5Cu00fcller-R%5Cu00f6ver%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22V.%20A.%22%2C%22lastName%22%3A%22Botchkarev%22%7D%5D%2C%22abstractNote%22%3A%22The%20hair%20follicle%20%28HF%29%20is%20the%20only%20mammalian%20organ%20that%20undergoes%20life-long%2C%20cyclic%20transformations%20from%20long%20stages%20of%20growth%20%28anagen%29%2C%20via%20rapid%2C%20apoptosis-driven%20organ%20involution%20%28catagen%29%20to%20a%20stage%20of%20relative%20%26quot%3Bresting%26quot%3B%20%28telogen%29.%20The%20controls%20that%20underlie%20these%20transformations%20clearly%20reside%20in%20and%5C%2For%20around%20the%20HF%20itself%2C%20and%20are%20likely%20to%20reflect%20-%20essentially%20autonomous%2C%20yet%20highly%20manipulable%20-%20changes%20in%20the%20local%20signalling%20milieu%20of%20e.g.%2C%20hair%20growth-modulatory%20growth%20factors%2C%20cytokines%2C%20hormones%20and%20adhesion%20molecules.%20Yet%20the%20molecular%20nature%20and%20organization%20of%20the%20%26quot%3Bhair%20cycle%20clock%26quot%3B%20%28HCC%29%20that%20drives%20these%20cyclic%20switches%20in%20the%20local%20signalling%20milieu%20remain%20obscure%2C%20and%20there%20is%20not%20even%20a%20fully%20satisfactory%20theory%20of%20hair%20cycle%20control.%20Since%20deciphering%20of%20the%20HCC%20is%20of%20paramount%20clinical%20importance%2C%20and%20since%20corresponding%20working%20hypotheses%20are%20badly%20needed%20to%20guide%20the%20design%20of%20more%20incisive%20experiments%20that%20identify%20the%20elusive%20central%20%26quot%3Boscillator%26quot%3B%20mechanism%20behind%20the%20HCC%2C%20we%20discuss%20basic%20requirements%20any%20convincing%20HCC%20theory%20should%20meet.%20After%20arguing%20that%20at%20least%20four%20distinct%20timing%20devices%20underlie%20HF%20chronobiology%20%28%26quot%3Bmorphogenesis%20clock%26quot%3B%2C%20%26quot%3Bcycling%20inducer%26quot%3B%2C%20%26quot%3Bdesynchronizer%26quot%3B%2C%20and%20the%20actual%20HCC%29%2C%20previously%20proposed%20HCC%20theories%20are%20briefly%20and%20critically%20reviewed.%20In%20the%20light%20of%20intriguing%20regulatory%20similarities%20between%20the%20HCC%20and%20the%20cell%20cycle%20machinery%2C%20we%20suggest%20here%20that%20the%20HCC%20may%20be%20driven%20by%20autonomous%2C%20cell%20cycle-coupled%20secretory%20activities%20of%20the%20HF%20mesenchyme%2C%20namely%20by%20changes%20in%20the%20G0%5C%2FG1-associated%20secretion%20of%20%26quot%3Bpapilla%20morphogens%26quot%3B%20by%20dermal%20papilla%20fibroblasts.%20Hopefully%2C%20this%20provocative%20hypothesis%20will%20encourage%20the%20proposition%20of%20novel%2C%20comprehensive%20HCC%20theories.%22%2C%22date%22%3A%221999-12%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1038%5C%2Fsj.jidsp.5640241%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%221087-0024%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A21%3A50Z%22%7D%7D%2C%7B%22key%22%3A%22SP5A8BKW%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Pi%5Cu00e9rard-Franchimont%20and%20Pi%5Cu00e9rard%22%2C%22parsedDate%22%3A%222001%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BPi%26%23xE9%3Brard-Franchimont%20C%2C%20Pi%26%23xE9%3Brard%20GE.%20Teloptosis%2C%20a%20turning%20point%20in%20hair%20shedding%20biorhythms.%20Dermatology.%202001%3B203%282%29%3A115%26%23x2013%3B7.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Teloptosis%2C%20a%20turning%20point%20in%20hair%20shedding%20biorhythms%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22C.%22%2C%22lastName%22%3A%22Pi%5Cu00e9rard-Franchimont%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22G.%20E.%22%2C%22lastName%22%3A%22Pi%5Cu00e9rard%22%7D%5D%2C%22abstractNote%22%3A%22The%20timing%20regulation%20of%20the%20hair%20growth%20phases%20is%20quite%20complex.%20We%20name%20teloptosis%20the%20termination%20of%20the%20telogen%20phase%20with%20hair%20shedding.%20The%20initiation%20of%20the%20next%20anagen%20I%20phase%20is%20independent%20from%20teloptosis.%20Both%20events%20are%20triggered%20by%20distinct%20factors.%20Indeed%2C%20teloptosis%20is%20the%20result%20of%20the%20loss%20of%20adhesion%20between%20cells%20of%20the%20club%20hair%20and%20those%20of%20its%20epithelial%20envelope.%20Teloptosis%20synchronization%20is%20perceived%20as%20telogen%20effluvium%2C%20related%20or%20not%20to%20shortening%20of%20the%20anagen%20phase.%20Subtle%20changes%20leading%20to%20early%20or%20delayed%20teloptosis%20affect%20hair%20density.%20It%20is%20possible%20to%20distinguish%20two%20types%20of%20telogen%20hairs%20on%20a%20trichogram.%20Those%20with%20an%20epithelial%20sheath%20indicate%20strong%20binding%2C%20suggesting%20early%20telogen%2C%20whereas%20the%20absence%20of%20an%20epithelial%20sheath%20indicates%20loose%20binding%2C%20conveying%20teloptosis.%22%2C%22date%22%3A%222001%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1159%5C%2F000051723%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%221018-8665%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A36%3A45Z%22%7D%7D%2C%7B%22key%22%3A%22BIKNPAS9%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Stenn%20and%20Paus%22%2C%22parsedDate%22%3A%222001-01%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BStenn%20KS%2C%20Paus%20R.%20Controls%20of%20hair%20follicle%20cycling.%20Physiol%20Rev.%202001%20Jan%3B81%281%29%3A449%26%23x2013%3B94.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Controls%20of%20hair%20follicle%20cycling%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22K.%20S.%22%2C%22lastName%22%3A%22Stenn%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22R.%22%2C%22lastName%22%3A%22Paus%22%7D%5D%2C%22abstractNote%22%3A%22Nearly%2050%20years%20ago%2C%20Chase%20published%20a%20review%20of%20hair%20cycling%20in%20which%20he%20detailed%20hair%20growth%20in%20the%20mouse%20and%20integrated%20hair%20biology%20with%20the%20biology%20of%20his%20day.%20In%20this%20review%20we%20have%20used%20Chase%20as%20our%20model%20and%20tried%20to%20put%20the%20adult%20hair%20follicle%20growth%20cycle%20in%20perspective.%20We%20have%20tried%20to%20sketch%20the%20adult%20hair%20follicle%20cycle%2C%20as%20we%20know%20it%20today%20and%20what%20needs%20to%20be%20known.%20Above%20all%2C%20we%20hope%20that%20this%20work%20will%20serve%20as%20an%20introduction%20to%20basic%20biologists%20who%20are%20looking%20for%20a%20defined%20biological%20system%20that%20illustrates%20many%20of%20the%20challenges%20of%20modern%20biology%3A%20cell%20differentiation%2C%20epithelial-mesenchymal%20interactions%2C%20stem%20cell%20biology%2C%20pattern%20formation%2C%20apoptosis%2C%20cell%20and%20organ%20growth%20cycles%2C%20and%20pigmentation.%20The%20most%20important%20theme%20in%20studying%20the%20cycling%20hair%20follicle%20is%20that%20the%20follicle%20is%20a%20regenerating%20system.%20By%20traversing%20the%20phases%20of%20the%20cycle%20%28growth%2C%20regression%2C%20resting%2C%20shedding%2C%20then%20growth%20again%29%2C%20the%20follicle%20demonstrates%20the%20unusual%20ability%20to%20completely%20regenerate%20itself.%20The%20basis%20for%20this%20regeneration%20rests%20in%20the%20unique%20follicular%20epithelial%20and%20mesenchymal%20components%20and%20their%20interactions.%20Recently%2C%20some%20of%20the%20molecular%20signals%20making%20up%20these%20interactions%20have%20been%20defined.%20They%20involve%20gene%20families%20also%20found%20in%20other%20regenerating%20systems%20such%20as%20fibroblast%20growth%20factor%2C%20transforming%20growth%20factor-beta%2C%20Wnt%20pathway%2C%20Sonic%20hedgehog%2C%20neurotrophins%2C%20and%20homeobox.%20For%20the%20immediate%20future%2C%20our%20challenge%20is%20to%20define%20the%20molecular%20basis%20for%20hair%20follicle%20growth%20control%2C%20to%20regenerate%20a%20mature%20hair%20follicle%20in%20vitro%20from%20defined%20populations%2C%20and%20to%20offer%20real%20solutions%20to%20our%20patients%26%23039%3B%20problems.%22%2C%22date%22%3A%222001-01%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1152%5C%2Fphysrev.2001.81.1.449%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%220031-9333%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A37%3A40Z%22%7D%7D%2C%7B%22key%22%3A%226CR2WRWS%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Rebora%20and%20Guarrera%22%2C%22parsedDate%22%3A%222002%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BRebora%20A%2C%20Guarrera%20M.%20Kenogen.%20A%20new%20phase%20of%20the%20hair%20cycle%3F%20Dermatology.%202002%3B205%282%29%3A108%26%23x2013%3B10.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Kenogen.%20A%20new%20phase%20of%20the%20hair%20cycle%3F%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Alfredo%22%2C%22lastName%22%3A%22Rebora%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Marcella%22%2C%22lastName%22%3A%22Guarrera%22%7D%5D%2C%22abstractNote%22%3A%22BACKGROUND%3A%20A%20novel%20phenomenon%20has%20been%20described%20by%20the%20phototrichogram%3A%20the%20emptiness%20of%20the%20follicle%20after%20teloptosis.%20We%20called%20this%20phenomenon%20kenogen%2C%20from%20the%20Greek%20kappaepsilonnu%5Cu00f3varsigma%2C%20%26%23039%3Bempty%26%23039%3B.%5CnOBJECTIVE%3A%20To%20describe%20the%20kenogen%20phase%20in%20its%20details.%5CnMETHODS%3A%20Analysis%20of%20the%20existing%20literature.%5CnRESULTS%3A%20The%20original%20observation%20in%202%20women%20was%20confirmed%20in%2010%20balding%20and%20non-balding%20males%20studied%20for%2014%20years%20in%20whom%20kenogen%20lasted%20about%204%20months%20increasing%20up%20to%20about%207%20months%20and%20affecting%2080%25%20of%20all%20hair%20cycles.%20In%202%20women%20with%20progressing%20androgenetic%20alopecia%20studied%20for%202%20years%2C%20kenogen%20involved%2022%25%20of%20the%20hair%20follicles%2C%20lasting%20from%203%20months%20to%201%20year.%20In%20a%20prepubertal%20boy%20studied%20for%201%20year%2C%20it%20involved%208%25%20of%20hairs%20and%20lasted%20about%202%20months.%5CnCONCLUSION%3A%20During%20kenogen%2C%20the%20hair%20follicle%20rests%20physiologically%2C%20but%20duration%20and%20frequency%20are%20greater%20in%20androgenetic%20alopecia%2C%20possibly%20accounting%20for%20baldness.%20In%20addition%20to%20the%20classical%20cycle%2C%20the%20hair%20follicle%20may%20follow%20an%20alternative%20route%20during%20which%20the%20telogen%20phase%2C%20not%20accompanied%20by%20a%20coincident%20new%20early%20anagen%2C%20ends%20with%20teloptosis%20leaving%20the%20follicle%20empty.%22%2C%22date%22%3A%222002%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1159%5C%2F000063908%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%221018-8665%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A35%3A18Z%22%7D%7D%2C%7B%22key%22%3A%22EMR7Y9HA%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Schneider%20et%20al.%22%2C%22parsedDate%22%3A%222009-02-10%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BSchneider%20MR%2C%20Schmidt-Ullrich%20R%2C%20Paus%20R.%20The%20hair%20follicle%20as%20a%20dynamic%20miniorgan.%20Curr%20Biol.%202009%20Feb%2010%3B19%283%29%3AR132-142.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22The%20hair%20follicle%20as%20a%20dynamic%20miniorgan%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Marlon%20R.%22%2C%22lastName%22%3A%22Schneider%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Ruth%22%2C%22lastName%22%3A%22Schmidt-Ullrich%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Ralf%22%2C%22lastName%22%3A%22Paus%22%7D%5D%2C%22abstractNote%22%3A%22Hair%20is%20a%20primary%20characteristic%20of%20mammals%2C%20and%20exerts%20a%20wide%20range%20of%20functions%20including%20thermoregulation%2C%20physical%20protection%2C%20sensory%20activity%2C%20and%20social%20interactions.%20The%20hair%20shaft%20consists%20of%20terminally%20differentiated%20keratinocytes%20that%20are%20produced%20by%20the%20hair%20follicle.%20Hair%20follicle%20development%20takes%20place%20during%20fetal%20skin%20development%20and%20relies%20on%20tightly%20regulated%20ectodermal-mesodermal%20interactions.%20After%20birth%2C%20mature%20and%20actively%20growing%20hair%20follicles%20eventually%20become%20anchored%20in%20the%20subcutis%2C%20and%20periodically%20regenerate%20by%20spontaneously%20undergoing%20repetitive%20cycles%20of%20growth%20%28anagen%29%2C%20apoptosis-driven%20regression%20%28catagen%29%2C%20and%20relative%20quiescence%20%28telogen%29.%20Our%20molecular%20understanding%20of%20hair%20follicle%20biology%20relies%20heavily%20on%20mouse%20mutants%20with%20abnormalities%20in%20hair%20structure%2C%20growth%2C%20and%5C%2For%20pigmentation.%20These%20mice%20have%20allowed%20novel%20insights%20into%20important%20general%20molecular%20and%20cellular%20processes%20beyond%20skin%20and%20hair%20biology%2C%20ranging%20from%20organ%20induction%2C%20morphogenesis%20and%20regeneration%2C%20to%20pigment%20and%20stem%20cell%20biology%2C%20cell%20proliferation%2C%20migration%20and%20apoptosis.%20In%20this%20review%2C%20we%20present%20basic%20concepts%20of%20hair%20follicle%20biology%20and%20summarize%20important%20recent%20advances%20in%20the%20field.%22%2C%22date%22%3A%222009-02-10%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1016%5C%2Fj.cub.2008.12.005%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%221879-0445%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A37%3A56Z%22%7D%7D%2C%7B%22key%22%3A%22I2Z4MZNU%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Higgins%20et%20al.%22%2C%22parsedDate%22%3A%222009-09%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BHiggins%20CA%2C%20Westgate%20GE%2C%20Jahoda%20CAB.%20From%20telogen%20to%20exogen%3A%20mechanisms%20underlying%20formation%20and%20subsequent%20loss%20of%20the%20hair%20club%20fiber.%20J%20Invest%20Dermatol.%202009%20Sept%3B129%289%29%3A2100%26%23x2013%3B8.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22From%20telogen%20to%20exogen%3A%20mechanisms%20underlying%20formation%20and%20subsequent%20loss%20of%20the%20hair%20club%20fiber%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Claire%20A.%22%2C%22lastName%22%3A%22Higgins%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Gillian%20E.%22%2C%22lastName%22%3A%22Westgate%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Colin%20A.%20B.%22%2C%22lastName%22%3A%22Jahoda%22%7D%5D%2C%22abstractNote%22%3A%22The%20hair%20follicle%20has%20the%20unique%20capacity%20to%20undergo%20periods%20of%20growth%2C%20regression%2C%20and%20rest%20before%20regenerating%20itself%20to%20restart%20the%20cycle.%20This%20dynamic%20cycling%20capacity%20enables%20mammals%20to%20change%20their%20coats%2C%20and%20for%20hair%20length%20to%20be%20controlled%20on%20different%20body%20sites.%20More%20recently%2C%20the%20process%20of%20club%20fiber%20shedding%20has%20been%20described%20as%20a%20distinct%20cycle%20phase%20known%20as%20exogen%2C%20and%20proposed%20to%20be%20an%20active%20phase%20of%20the%20hair%20cycle.%20This%20review%20focuses%20on%20the%20importance%20of%20the%20shedding%20phase%20of%20the%20hair%20cycle%20and%2C%20in%20the%20context%20of%20current%20literature%2C%20analyzes%20the%20processes%20of%20club%20fiber%20formation%2C%20retention%2C%20and%20release%2C%20which%20may%20influence%20progression%20through%20exogen%2C%20particularly%20in%20relation%20to%20human%20hair.%22%2C%22date%22%3A%222009-09%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1038%5C%2Fjid.2009.66%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%221523-1747%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A33%3A56Z%22%7D%7D%2C%7B%22key%22%3A%224WMX33EI%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Buffoli%20et%20al.%22%2C%22parsedDate%22%3A%222014-03%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BBuffoli%20B%2C%20Rinaldi%20F%2C%20Labanca%20M%2C%20Sorbellini%20E%2C%20Trink%20A%2C%20Guanziroli%20E%2C%20et%20al.%20The%20human%20hair%3A%20from%20anatomy%20to%20physiology.%20Int%20J%20Dermatol.%202014%20Mar%3B53%283%29%3A331%26%23x2013%3B41.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22The%20human%20hair%3A%20from%20anatomy%20to%20physiology%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Barbara%22%2C%22lastName%22%3A%22Buffoli%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Fabio%22%2C%22lastName%22%3A%22Rinaldi%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Mauro%22%2C%22lastName%22%3A%22Labanca%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Elisabetta%22%2C%22lastName%22%3A%22Sorbellini%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Anna%22%2C%22lastName%22%3A%22Trink%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Elena%22%2C%22lastName%22%3A%22Guanziroli%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Rita%22%2C%22lastName%22%3A%22Rezzani%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Luigi%20F.%22%2C%22lastName%22%3A%22Rodella%22%7D%5D%2C%22abstractNote%22%3A%22BACKGROUND%3A%20Hair%20is%20a%20unique%20character%20of%20mammals%20and%20has%20several%20functions%2C%20from%20protection%20of%20the%20skin%20to%20sexual%20and%20social%20communication.%20In%20literature%2C%20there%20are%20various%20studies%20about%20hair%20that%20take%20into%20consideration%20different%20aspects%20within%20many%20fields%20of%20science%2C%20including%20biology%2C%20dermatology%2C%20cosmetics%2C%20forensic%20sciences%2C%20and%20medicine.%5CnMETHODS%3A%20We%20carried%20out%20a%20search%20of%20studies%20published%20in%20PubMed%20up%20to%202013.%5CnRESULTS%3A%20In%20this%20review%2C%20we%20summarized%20the%20principal%20anatomical%20and%20physiological%20aspects%20of%20the%20different%20types%20of%20human%20hair%2C%20and%20we%20considered%20the%20clinical%20significance%20of%20the%20different%20structures%20and%20the%20distribution%20of%20the%20hair%20in%20the%20human%20body.%5CnCONCLUSION%3A%20This%20review%20could%20be%20the%20basis%20for%20improvement%20and%20progression%20in%20the%20field%20of%20hair%20research.%22%2C%22date%22%3A%222014-03%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1111%5C%2Fijd.12362%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%221365-4632%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A38%3A24Z%22%7D%7D%2C%7B%22key%22%3A%22I93UBY9X%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Oh%20et%20al.%22%2C%22parsedDate%22%3A%222016-01%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BOh%20JW%2C%20Kloepper%20J%2C%20Langan%20EA%2C%20Kim%20Y%2C%20Yeo%20J%2C%20Kim%20MJ%2C%20et%20al.%20A%20Guide%20to%20Studying%20Human%20Hair%20Follicle%20Cycling%20In%20Vivo.%20J%20Invest%20Dermatol.%202016%20Jan%3B136%281%29%3A34%26%23x2013%3B44.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22A%20Guide%20to%20Studying%20Human%20Hair%20Follicle%20Cycling%20In%20Vivo%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Ji%20Won%22%2C%22lastName%22%3A%22Oh%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Jennifer%22%2C%22lastName%22%3A%22Kloepper%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Ewan%20A.%22%2C%22lastName%22%3A%22Langan%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Yongsoo%22%2C%22lastName%22%3A%22Kim%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Joongyeub%22%2C%22lastName%22%3A%22Yeo%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Min%20Ji%22%2C%22lastName%22%3A%22Kim%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Tsai-Ching%22%2C%22lastName%22%3A%22Hsi%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Christian%22%2C%22lastName%22%3A%22Rose%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Ghil%20Suk%22%2C%22lastName%22%3A%22Yoon%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Seok-Jong%22%2C%22lastName%22%3A%22Lee%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22John%22%2C%22lastName%22%3A%22Seykora%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Jung%20Chul%22%2C%22lastName%22%3A%22Kim%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Young%20Kwan%22%2C%22lastName%22%3A%22Sung%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Moonkyu%22%2C%22lastName%22%3A%22Kim%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Ralf%22%2C%22lastName%22%3A%22Paus%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Maksim%20V.%22%2C%22lastName%22%3A%22Plikus%22%7D%5D%2C%22abstractNote%22%3A%22Hair%20follicles%20%28HFs%29%20undergo%20lifelong%20cyclical%20transformations%2C%20progressing%20through%20stages%20of%20rapid%20growth%20%28anagen%29%2C%20regression%20%28catagen%29%2C%20and%20relative%20%26quot%3Bquiescence%26quot%3B%20%28telogen%29.%20Given%20that%20HF%20cycling%20abnormalities%20underlie%20many%20human%20hair%20growth%20disorders%2C%20the%20accurate%20classification%20of%20individual%20cycle%20stages%20within%20skin%20biopsies%20is%20clinically%20important%20and%20essential%20for%20hair%20research.%20For%20preclinical%20human%20hair%20research%20purposes%2C%20human%20scalp%20skin%20can%20be%20xenografted%20onto%20immunocompromised%20mice%20to%20study%20human%20HF%20cycling%20and%20manipulate%20long-lasting%20anagen%20in%20vivo.%20Although%20available%20for%20mice%2C%20a%20comprehensive%20guide%20on%20how%20to%20recognize%20different%20human%20hair%20cycle%20stages%20in%20vivo%20is%20lacking.%20In%20this%20article%2C%20we%20present%20such%20a%20guide%2C%20which%20uses%20objective%2C%20well-defined%2C%20and%20reproducible%20criteria%2C%20and%20integrates%20simple%20morphological%20indicators%20with%20advanced%2C%20%28immuno%29-histochemical%20markers.%20This%20guide%20also%20characterizes%20human%20HF%20cycling%20in%20xenografts%20and%20highlights%20the%20utility%20of%20this%20model%20for%20in%20vivo%20hair%20research.%20Detailed%20schematic%20drawings%20and%20representative%20micrographs%20provide%20examples%20of%20how%20best%20to%20identify%20human%20HF%20stages%2C%20even%20in%20suboptimally%20sectioned%20tissue%2C%20and%20practical%20recommendations%20are%20given%20for%20designing%20human-on-mouse%20hair%20cycle%20experiments.%20Thus%2C%20this%20guide%20seeks%20to%20offer%20a%20benchmark%20for%20human%20hair%20cycle%20stage%20classification%2C%20for%20both%20hair%20research%20experts%20and%20newcomers%20to%20the%20field.%22%2C%22date%22%3A%222016-01%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.1038%5C%2FJID.2015.354%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%221523-1747%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A22%3A59Z%22%7D%7D%2C%7B%22key%22%3A%22PFFVSYJG%22%2C%22library%22%3A%7B%22id%22%3A11711645%7D%2C%22meta%22%3A%7B%22creatorSummary%22%3A%22Niu%20et%20al.%22%2C%22parsedDate%22%3A%222023-07-03%22%2C%22numChildren%22%3A0%7D%2C%22bib%22%3A%22%26lt%3Bdiv%20class%3D%26quot%3Bcsl-bib-body%26quot%3B%20style%3D%26quot%3Bline-height%3A%201.35%3B%20%26quot%3B%26gt%3B%5Cn%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-entry%26quot%3B%20style%3D%26quot%3Bclear%3A%20left%3B%20%26quot%3B%26gt%3B%5Cn%20%20%20%20%26lt%3Bdiv%20class%3D%26quot%3Bcsl-left-margin%26quot%3B%20style%3D%26quot%3Bfloat%3A%20left%3B%20padding-right%3A%200.5em%3B%20text-align%3A%20right%3B%20width%3A%201em%3B%26quot%3B%26gt%3B1.%26lt%3B%5C%2Fdiv%26gt%3B%26lt%3Bdiv%20class%3D%26quot%3Bcsl-right-inline%26quot%3B%20style%3D%26quot%3Bmargin%3A%200%20.4em%200%201.5em%3B%26quot%3B%26gt%3BNiu%20Y%2C%20Wang%20Y%2C%20Chen%20H%2C%20Liu%20X%2C%20Liu%20J.%20Overview%20of%20the%20Circadian%20Clock%20in%20the%20Hair%20Follicle%20Cycle.%20Biomolecules.%202023%20July%203%3B13%287%29%3A1068.%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%20%20%20%26lt%3B%5C%2Fdiv%26gt%3B%5Cn%26lt%3B%5C%2Fdiv%26gt%3B%22%2C%22data%22%3A%7B%22itemType%22%3A%22journalArticle%22%2C%22title%22%3A%22Overview%20of%20the%20Circadian%20Clock%20in%20the%20Hair%20Follicle%20Cycle%22%2C%22creators%22%3A%5B%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Ye%22%2C%22lastName%22%3A%22Niu%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Yujie%22%2C%22lastName%22%3A%22Wang%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Hao%22%2C%22lastName%22%3A%22Chen%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Xiaomei%22%2C%22lastName%22%3A%22Liu%22%7D%2C%7B%22creatorType%22%3A%22author%22%2C%22firstName%22%3A%22Jinyu%22%2C%22lastName%22%3A%22Liu%22%7D%5D%2C%22abstractNote%22%3A%22The%20circadian%20clock%20adapts%20to%20the%20light-dark%20cycle%20and%20autonomously%20generates%20physiological%20and%20metabolic%20rhythmicity.%20Its%20activity%20depends%20on%20the%20central%20suprachiasmatic%20pacemaker.%20However%2C%20it%20also%20has%20an%20independent%20function%20in%20peripheral%20tissues%20such%20as%20the%20liver%2C%20adipose%20tissue%2C%20and%20skin%2C%20which%20integrate%20environmental%20signals%20and%20energy%20homeostasis.%20Hair%20follicles%20%28HFs%29%20maintain%20homeostasis%20through%20the%20HF%20cycle%2C%20which%20depends%20heavily%20on%20HF%20stem%20cell%20self-renewal%20and%20the%20related%20metabolic%20reprogramming.%20Studies%20have%20shown%20that%20circadian%20clock%20dysregulation%20in%20HFs%20perturbs%20cell%20cycle%20progression.%20Moreover%2C%20there%20is%20increasing%20evidence%20that%20the%20circadian%20clock%20exerts%20a%20significant%20influence%20on%20glucose%20metabolism%2C%20feeding%5C%2Ffasting%2C%20stem%20cell%20differentiation%2C%20and%20senescence.%20This%20suggests%20that%20circadian%20metabolic%20crosstalk%20plays%20an%20essential%20role%20in%20regulating%20HF%20regeneration.%20An%20improved%20understanding%20of%20the%20role%20of%20the%20circadian%20clock%20in%20HFs%20may%20facilitate%20the%20discovery%20of%20new%20drug%20targets%20for%20hair%20loss.%20Therefore%2C%20the%20present%20review%20provides%20a%20discussion%20of%20the%20relationship%20between%20the%20circadian%20clock%20and%20HF%20regeneration%2C%20mainly%20from%20the%20perspective%20of%20HF%20metabolism%2C%20and%20summarizes%20the%20current%20understanding%20of%20the%20mechanisms%20by%20which%20HFs%20function.%22%2C%22date%22%3A%222023-07-03%22%2C%22section%22%3A%22%22%2C%22partNumber%22%3A%22%22%2C%22partTitle%22%3A%22%22%2C%22DOI%22%3A%2210.3390%5C%2Fbiom13071068%22%2C%22citationKey%22%3A%22%22%2C%22url%22%3A%22%22%2C%22PMID%22%3A%22%22%2C%22PMCID%22%3A%22%22%2C%22ISSN%22%3A%222218-273X%22%2C%22language%22%3A%22eng%22%2C%22collections%22%3A%5B%22DWTP59MQ%22%5D%2C%22dateModified%22%3A%222025-07-28T15%3A34%3A38Z%22%7D%7D%5D%7D 1.
Chase HB, Montagna W, Malone JD. Changes in the skin in relation to the hair growth cycle. Anat Rec. 1953 May;116(1):75–81.
1.
Chase HB. Growth of the hair. Physiol Rev. 1954 Jan;34(1):113–26.
1.
Van Scott EJ, Reinertson RP, Steinmuller R. The growing hair roots of the human scalp and morphologic changes therein following amethopterin therapy. J Invest Dermatol. 1957 Sept;29(3):197–204.
1.
Pelfini C, Cerimele D, Pisanu G. Aging of the skin and hair growth in man. In: Hair growth. London: Pergamon Press; 1969. p. 153–60.
1.
Saitoh M, Uzuka M, Sakamoto M. Human hair cycle. J Invest Dermatol. 1970 Jan;54(1):65–81.
1.
Moretti G, Rampini E, Rebora A. The hair cycle re-evaluated. Int J Dermatol. 1976 May;15(4):277–85.
1.
Ebling FJ. The hair cycle and its regulation. Clin Dermatol. 1988;6(4):67–73.
1.
Kligman AM. The comparative histopathology of male-pattern baldness and senescent baldness. Clin Dermatol. 1988;6(4):108–18.
1.
Courtois M, Loussouarn G, Hourseau C, Grollier JF. Hair cycle and alopecia. Skin Pharmacol. 1994;7(1–2):84–9.
1.
Paus R, Cotsarelis G. The biology of hair follicles. N Engl J Med. 1999 Aug 12;341(7):491–7.
1.
Paus R, Müller-Röver S, Botchkarev VA. Chronobiology of the hair follicle: hunting the “ hair cycle clock.” J Investig Dermatol Symp Proc. 1999 Dec;4(3):338–45.
1.
Piérard-Franchimont C, Piérard GE. Teloptosis, a turning point in hair shedding biorhythms. Dermatology. 2001;203(2):115–7.
1.
Stenn KS, Paus R. Controls of hair follicle cycling. Physiol Rev. 2001 Jan;81(1):449–94.
1.
Rebora A, Guarrera M. Kenogen. A new phase of the hair cycle? Dermatology. 2002;205(2):108–10.
1.
Schneider MR, Schmidt-Ullrich R, Paus R. The hair follicle as a dynamic miniorgan. Curr Biol. 2009 Feb 10;19(3):R132-142.
1.
Higgins CA, Westgate GE, Jahoda CAB. From telogen to exogen: mechanisms underlying formation and subsequent loss of the hair club fiber. J Invest Dermatol. 2009 Sept;129(9):2100–8.
1.
Buffoli B, Rinaldi F, Labanca M, Sorbellini E, Trink A, Guanziroli E, et al. The human hair: from anatomy to physiology. Int J Dermatol. 2014 Mar;53(3):331–41.
1.
Oh JW, Kloepper J, Langan EA, Kim Y, Yeo J, Kim MJ, et al. A Guide to Studying Human Hair Follicle Cycling In Vivo. J Invest Dermatol. 2016 Jan;136(1):34–44.
1.
Niu Y, Wang Y, Chen H, Liu X, Liu J. Overview of the Circadian Clock in the Hair Follicle Cycle. Biomolecules. 2023 July 3;13(7):1068.